[白川一] 分化專一化的分歧路徑:保衛細胞、芥子酶細胞及其延伸
發稿時間: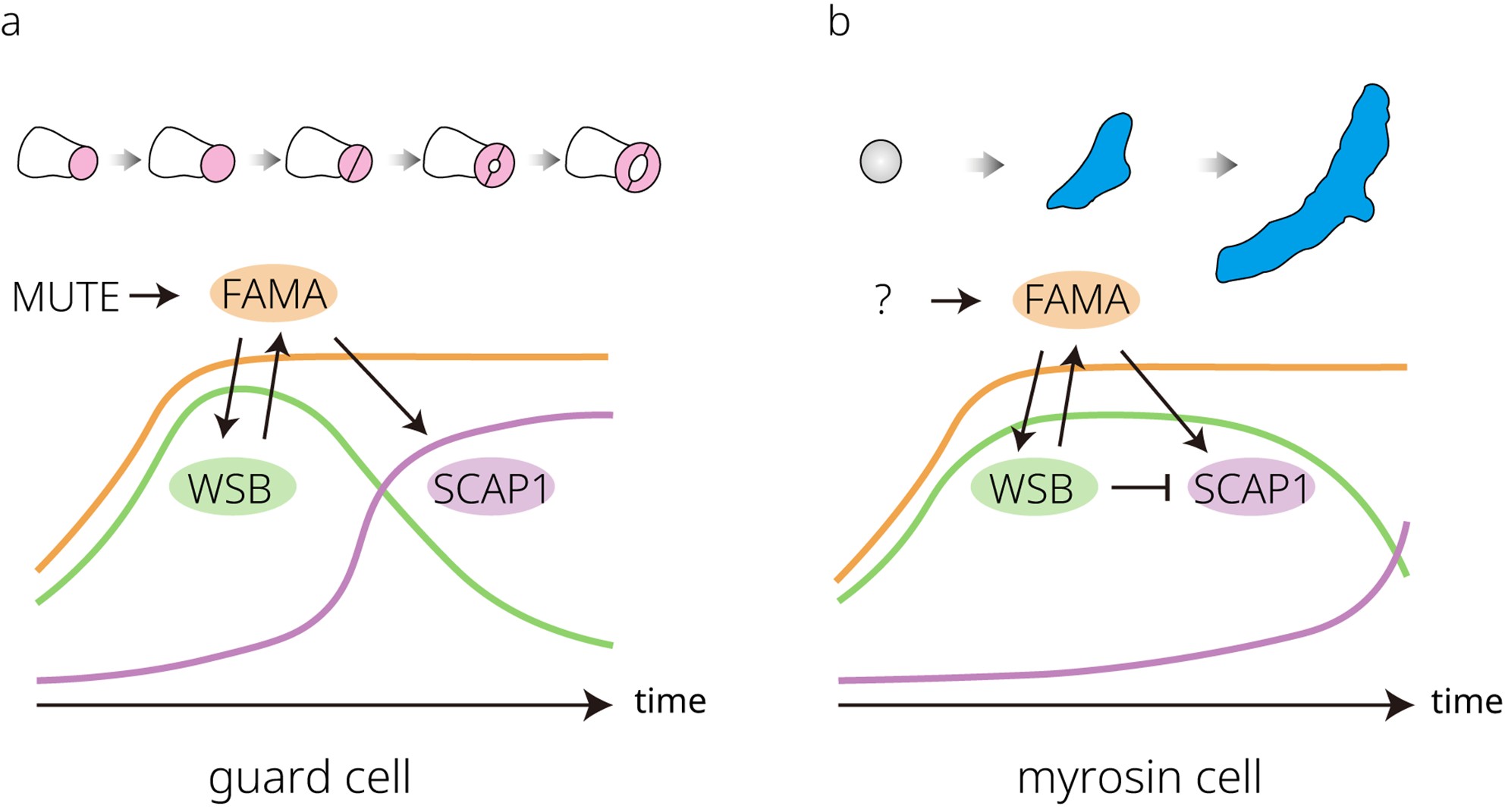
保衛細胞與芥子酶細胞的發育軌跡與調控動態 (a) 在氣孔譜系細胞中,bHLH 轉錄因子 MUTE 可直接誘導 FAMA 的表現。FAMA 轉譯後進一步調控 WASABI MAKER(WSB)與 STOMATAL CARPENTER 1(SCAP1)的表現。在氣孔譜系中,WSB 的表現早於 SCAP1,且表現時間窗較窄;然而兩者的表現時間窗部分重疊。SCAP1 的表現於較晚期上升並在保衛細胞(GCs)中持續存在,而其在成熟保衛細胞中對氣孔運動所必需。上述因子與 FAMA 共同協調保衛細胞分化的最後步驟。 (b) 芥子酶細胞由基本分生組織(ground meristem)細胞直接分化而來。這些細胞中 FAMA 的表現被啟動,並促進 WSB 的表現。相較於在保衛細胞中的表現,WSB 在芥子酶細胞中具有較寬廣的表現時間窗,並可透過回饋迴路維持 FAMA 的表現。在分化的大部分期間,SCAP1 的表現受到 WSB 抑制,但在成熟末期變得可偵測,可能參與芥子酶細胞分化的最後步驟。
本篇綜述探討植物如何透過重複利用保守性的轉錄因子,而非另行發明全新的調控系統,來產生新的專一化細胞類型。我們聚焦於轉錄因子 FAMA:其負責調控氣孔保衛細胞分化,並被共同選用(co-opted)用以生成屬於十字花目(Brassicales)特有的防禦細胞類型——芥子酶細胞。近期研究顯示,下游基因啟動在時間點與活化持續性上的差異,使同一調控因子得以指定不同的細胞命運。單細胞轉錄體學進一步在根部發現先前未預期的芥子酶細胞,突顯不同組織間的發育可塑性。綜合而言,這些發現說明既有基因網路在演化過程中經由重新接線(rewiring),為得以驅動植物細胞專一化的重要機制。